|
Nawet gdy mamy do czynienia z
drugim wydaniem podręcznika akademickiego, poprawionym i uzupełnionym,
zwykle nie wszystkie jego słabości zostały wcześniej zauważone przez autora,
recenzentów, korektę i redakcję. Te słabości widoczne są najlepiej przez
studentów i osoby spotykające się po raz pierwszy z tematyką podręcznika. Coś
co autorowi wydaje się oczywiste, dla czytelnika może być mętnym opisem
pełnym myślowych przeskoków. Dlatego bardzo proszę wszystkich czytelników o
przesyłanie wszelkich skarg i pretensji dotyczących tej książki na mój adres
e-mailowy (adam.lomnicki@uj.edu.pl).
Cenne dla mnie będą nie tylko znalezione błędy, ale także różnego rodzaju
niejasności. Czytelnikom, którzy do mnie napiszą postaram się odpowiedzieć i
wyjaśnić każdy fragment zbyt zawikłany, a potem tak zmienić książkę aby
niejasności usunąć. Ponieważ mamy już dwa wydania „Ekologii Ewolucyjnej”
poniżej znajdzie czytelnik niewielka erratę dla drugiego wydania i proponowane uzupełnienia do ewentualnych
dalszych wydań. Są tam też dwa zadania sprawdzające zrozumienie
pewnych fragmentów tekstu, które dotyczą obu wydań. Dla czytelników którzy dysponują jedynie
pierwszym wydaniem dodano poprzednią erratę i poprzednie
uzupełnienia, które zostały już wprowadzone do drugiego wydania. Errata do II wydania „Ekologii
Ewolucyjnej” na dzień 8 października 2013 roku Strona 215, wiersz 17 od góry,
w mianowniku lewej strony nierówności (D4.1): zamiast dN daj dNt UZUPEŁNIENIA do wprowadzenia przy trzecim
wydaniu Ekologii Ewolucyjnej Strona 122: po ostatnim wierszu na tej stronie
dodaj następujący nowy fragment i wprowadź odpowiednie zmiany w Piśmiennictwie: Genetyczne
podstawy utrzymywanie się zmienności w cechach epigenetycznych i koszt
utrzymywania tych cech opisany został
(Johnston et al. 2013) w oparciu o wieloletnie badania zdziczałych owiec na
niewielkiej wyspie (100 ha) Soay w archipelagu Hebryd Zewnętrznych znajdujących się na Atlantyku
północnym i należących do Szkocji.
Przypuszcza się, że owce te, stanowiące obecnie rasę owiec domowych (Ovis aries)
zostały tam wprowadzone przez człowieka ale zdziczały już 4 tysiące lat temu.
Są mniejsze od owcy hodowlanej i wyglądem przypominają muflony. Rogi u samców
tych owiec są określone przez jeden locus z dwoma
allelami. Allel
Ho+ daje u samców w homozygocie i w heterozygocie normalne
duże rogi, natomiast samce będace homozygotami allelu alternatywnego HoP
mają mniejsze rogi lub rogi szczątkowe. Ponieważ w latach 1985-2012 odłowiono
na wyspie Soay 5880 owiec określając ich genotypy i
zmienność genetyczną pozwalającą na ustalenie ojcostwa, autorzy mieli
możliwość określenia nie tylko przeżywalności dla 1343 samców, ale także
ich sukcesu reprodukcyjnego dla 640
osobników. Samce genotypu Ho+ Ho+ nie
różnią sukcesem reprodukcyjnym w sposób statystycznie istotny od samców -
heterozygot Ho+HoP,
natomiast samce genotypu HoP HoP mają znacznie niższy sukces
reprodukcyjny. Inaczej kształtuje się przeżywalność samców tych trzech
genotypów. Najlepiej przeżywają homozygoty HoP
HoP i heterozygoty Ho+HoP a różnice w
ich przeżywalności są statystycznie nieistotne, natomiast homozygoty Ho+
Ho+ przeżywają znacznie gorzej. Konsekwencją takich efektów
tych trzech genotypów jest najwyższe dostosowanie heterozygot Ho+HoP i znacznie
niższe dostosowanie obu homozygot Ho+ Ho+
oraz HoP HoP. Między samcami homozygotami nie ma w
dostosowaniu różnic statystycznie istotnych, istnieją natomiast takie różnice
między nimi a samcami heterozygotami Ho+HoP.
Mamy tu zatem do czynienia z naddominacją, ale
także z empirycznym dowodem, że utrzymywanie cech epigametycznych
dających sukcesy reprodukcyjne obniża inne elementy dostosowania, czyli jest
kosztowne. Zauważ, że przedstawione tu zależności są zależnościami
statystycznymi na które działa dobór naturalny. Natomiast pojedynczy samiec
może przeżywać nieco lepiej ze względu na lepszy zestaw genów w pozostałej
części jego genomu. Dlatego preferencja samic wobec samców z dużymi rogami
jest zrozumiała. Johnston S.
E., Gratten J., Berenos
C., Pilkiington J. G. Clutton-Brock
T.H, Pemberton J. M. i Slate J. 2013. Life
history trade-offs at a single locus maintain sexually selected genetic
variation. Nature 502: 93 – 96. Zadania
|
|
|
|
|
|||
|
Errata do pierwszego wydania
„Ekologii Ewolucyjnej” na dzień 5 marca 2013 roku Strona 4, wiersz 1 od góry:
zamiast Ilustracja na
okładce Piotr Ligier Muzeum Narodowe w Warszawie daj Ilustracja na okładce, to fragment obrazu Józefa
Chełmońskiego (1849-1914) pt. 'Kuropatwy na śniegu' (1891, olej na płótnie,
123x199 cm) ze zbiorów Muzeum Narodowego w Warszawie. Fotografia: Piotr Ligier, Muzeum Narodowe w Warszawie Strona 24, wiersz 3 i 4 od
góry: zamiast Samce można pominąć dlatego, że nie są zwykle czynnikiem
ograniczającym rozrodczość populacji daj W tym uproszczonym przykładzie założono, że
brak samców nie ogranicza rozrodczości samic, a przeżywalności samic i samców
są identyczne. W rzeczywistości te przeżywalności są różne a samce różnią się
miedzy sobą zdolnościami do zdobycia i zapłodnienia samicy. Dlatego w
bardziej realistycznych modelach dostosowanie powinno być oceniane osobno dla
każdej z płci. Strona 29, mianownik równania w
wierszu 13 od góry: zamiast M3 – M2 daj M2 – M1 Strona 35, wiersz 5 od góry:
zamiast Obie populacje daj Nasiona z obu populacji Strona 35, wiersz 9 od dołu (w podpisie
ryciny): zamiast Prawy wykres daj Lewy wykres Strona 48, wiersz 7 od góry: zamiast 10 minut daj 10
sekund Strona 48, wiersz 8 od góry: zamiast 50 minutach daj 50
sekundach, zamiast 100 minutach daj 100 sekundach Strona54,
wiersz 18 od góry: wykreśl słowa lub gorszą Strona
58, wiersz 17 od góry: zamiast Nierówność (4.8) daj
Nierówność (4.5) Strona 71, wiersz 2 od
dołu: zamiast (Hurst i in.) daj (Hurst i in. 1996) Strona 74, wiersz 15 i 14
od dołu: zamiast w
prawym górnym rogu symbol V daj symbol V
w prawej kolumnie drugiego wiersza Strona 75, wiersz 18 od
góry: zamiast
strategią ewolucyjnie stabilną daj strategią
wybieraną Strona 75, wiersz 21 od
góry: zamiast strategią
ewolucyjnie stabilną daj strategia wybieraną,
czyli że w obu przypadkach jest to strategia ewolucyjnie stabilna. Strona 75 wiersz 9 od
dołu: zamiast wówczas
strategią ewolucyjnie stabilną (ESS) jest daj wówczas
strategia wybierają jest Strona 75 wiersz 5 od
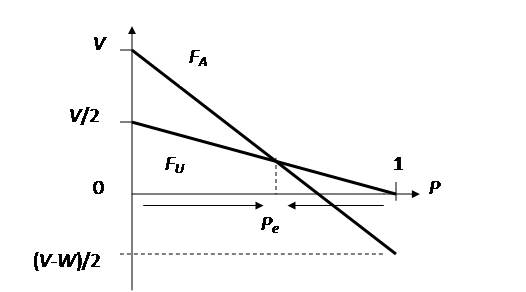
dołu: zamiast A jest ESS daj strategia wybieraną jest A Strona 77, rycina 5.1
powinna mieć zaznaczony symbol V na
osi pionowej a nie tylko V/2, tak
jak na poprawnej rycinie poniżej
Strona
82, wiersz 1 od góry w opisie ryciny 5.3: zamiast z ryciny 4.3 daj z ryciny 5.2 Strona
89, wiersz 2 od góry: zamiast FA i FB daj
FA i FU Strona 95, wiersze 7 do
10 od dołu: zdanie Najlepszy
dowód, że autor wspomnianej tu publikacji z roku 1981 – Robert Axelrod był doradcą Baracka Obamy w czasie wyborów
prezydenckich w Stanach Zjednoczonych w roku 2008. wykreśl,
ponieważ pomylono go z doradcą prezydenta B. Obamy - Davidem Axelrodem, jako że obaj są z wykształcenia politologami
(ang. political scientist),
a Robert nawet doradcą amerykańskiego Ministerstwa Obrony. Strona 125, wiersze 12,
13 i 14 od góry: zamiast a gdybyśmy rozważali tu
organizm, który może żyć znacznie dłużej, to z każdym rokiem różnica
byłaby jeszcze większa. daj a biorąc pod uwagę liczbę wnuków i prawnuków
osobników rozmnażających się wcześniej, różnica ta jest mniejsza, ale w
dalszym ciągu na korzyść osobników rozmnażających się później. Strona
145, wiersz 8 od dołu: zamiast Dodatku 2 daj Dodatku 4 Strona
147, wiersz 8 od góry: zamiast Dodatku 2 daj Dodatku 4 Strona
147, wiersz 11 od dołu: zamiast Dodatku 2 daj Dodatku 4 Strona
147, wiersz 10 od dołu: zamiast Dodatku 2 daj Dodatku 4 Strona
148, wiersz 17 od dołu: zamiast Dodatku 2 daj Dodatku 4 Strona 188, w tabeli D1,1
przy typie kojarzeń AA × aa częstości kojarzeń: zamiast 2PAAPAa daj 2PAAPaa Strona 190, tabela D1.1, czwarty wiersz z 6 w środku tabeli: zamiast AA × Aa daj Aa× Aa Strona 200, w ramce,
wiersz 10 od dołu: zamiast działanie doboru daj działanie dryfu Strona
210, wiersz 1 od dołu: zamiast Nt daj Nt+1 UZUPEŁNIENIA do pierwszego wydania Ekologii
Ewolucyjnej Strona 68: po wierszu 14 od
góry dodaj następujący nowy fragment i wprowadź odpowiednie zmiany w Piśmiennictwie: Opisany powyżej model teoretyczny
cykli drobnych gryzoni nie brał pod uwagę ich indywidualnych cech, szczegółów
ich behawioru, przestrzennego rozmieszczenia osobników i związanych z
tymi zjawiskami ich opóźnionych efektów na dynamikę populacji. Lemingi
i inne drobne gryzonie mogą jednak nie być najlepszym modelem do badań
ze względu na trudności w obserwacji ich zachowań
społecznych. Dlatego znaczenie doboru krewniaczego w powstawaniu cykli
populacyjnych zostało stwierdzone u łatwiejszych do obserwacji ptaków, a
ściślej u pardwy szkockiej (Lagopus lagopus scoticus). Model
teoretyczny (Hendry et al. 1997) i badania
empiryczne (Piertney et al. 2008) dotyczyły
dynamiki populacyjnej i zachowań społecznych
u terytorialnych samców tego gatunku, jako że zachowanie terytorialne
samców determinuje przebieg cyklu populacyjnego. Ponieważ tylko jedna samica
przebywająca z samcem na jego terytorium może wydawać potomstwo, zagęszczenie
populacji można mierzyć liczbą terytorialnych samców na jednostkę powierzchni
odpowiedniego dla nich siedliska. Model
teoretyczny zakładał, ze samce pardw rozpoznają jako krewniaków synów, ojców
i braci, czyli ze współczynnikiem pokrewieństwa r = 0,5. Gdy zagęszczenie par jest niewielkie wówczas tolerują
one terytoria krewniaków w sąsiedztwie swego terytorium i są gotowe ustąpić
nawet część swego terytorium osobnikom spokrewnionym. Prowadzi to do niskiej
agresji, wysokiej reprodukcji, powstawania wielu licznych skupisk osobników
spokrewnionych i wzrostu zagęszczenia całej populacji. Skupisko krewniacze
zostało zdefiniowane jako co najmniej dwa przylegające do siebie i należące do osobników spokrewnionych
terytoria. W wyniku wzrostu
zagęszczenia terytoria stają się coraz mniejsze i dalsze ustępowanie miejsca
nawet osobnikom spokrewnionym nie zwiększa dostosowania całkowitego (por.
str. 54), czyli innymi słowy straty z zachowania altruistycznego są znacznie
wyższe niż przy dużych terytoriach i nie spełniają nierówności (4.1).
Prowadzi to do agresji także wobec osobników spokrewnionych, które nie
znajdując miejsc obok terytoriów swoich krewniaków, przemieszczają się w
poszukiwaniu terytoriów dla siebie, co prowadzi do zaniku skupisk osobników
spokrewnionych. Powoduje to wzrost agresji miedzy samcami a po śmierci
właściciela terytorium, sąsiadujące osobniki nie pozwalają na osiedlenie się
nowego właściciela. W konsekwencji następuje
zmniejszenie liczby zajętych terytoriów, a tym samym zagęszczenia
populacji. Dopiero, gdy zagęszczenie samców spadnie tak, że nie dochodzi do
konfliktów między nimi, możliwy jest spadek agresji i ustępowanie miejsca w
sąsiedztwie osobnikom spokrewnionym. Procesy
zachodzące w populacjach pardwy mogą być szczegółowo śledzone przez indywidualne znakowanie każdego samca,
określenie jego terytorium oraz pobranie od niego próbki krwi dla określenia
mikro-satelitarnego DNA. Umożliwia to analizę pokrewieństwa miedzy samcami
oraz identyfikacje, położenie i liczbę skupisk osobników spokrewnionych.
Natężenie agresji między samcami mierzone było wysokością, tak zwanej przez polskich
myśliwych, róży czyli paska czerwonej narośli skórnej nad oczyma (ang. supra-orbital comb).
Było to możliwe, ponieważ wraz ze wzrostem agresywności samca rośnie szerokość
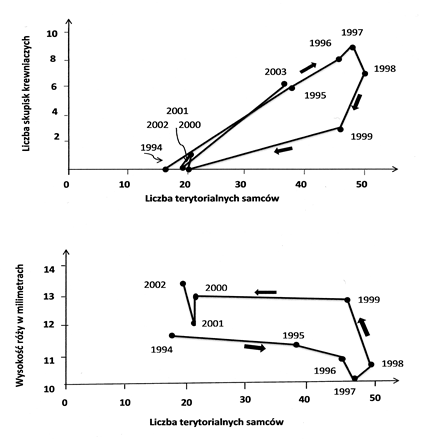
róży. Dane empiryczne przedstawione w pracy Piertneya
et al. (2008) były zebrane w latach 1994 ̶ 2003 na obszarze 52 hektarów
i są przedstawione na ryc. 4.4. Wyjaśniają one mechanizm powstawania cyklu.
Wzrost populacji rozpoczęty w roku 1994 przez 17 terytorialnych samców trwa
do roku 1998 osiągając liczbę 50 terytorialnych samców. Do roku 1997 wzrost ten idzie w parze
(górny wykres) ze wzrostem liczby
skupisk krewniaczych. Między
rokiem 1997 a 1998 spada liczba skupisk krewniaczych, a po roku 1998 spadek
tych skupisk jest coraz większy co idzie w parze ze spadkiem liczby
terytorialnych samców, poniżej 20 osobników. Drugą ważną cechą cyklu jest
natężenie agresji mierzone wysokością róży (dolny wykres). Agresja mierzona
wysokością róży jest bardzo mała w roku 1994
i maleje do wysokości róży wynoszącej 10 mm w roku 1997. Po tym roku
wraz ze spadkiem liczby skupisk rośnie wysokość róży osiągając w roku 1999
prawie 13 mm i przewyższając 13 mm w roku 2002. Gdyby liczba skupisk krewniaczych i poziom agresji
reagował natychmiast na zagęszczenie terytorialnych samców cykl populacyjny
byłby niemożliwy i populacja byłaby stabilna.
Tutaj oba procesy, a mianowicie tworzenie skupisk krewniaczych i
nasilanie się agresji, występują z pewnym opóźnieniem. Dorosłe samce
przeżywają rok z prawdopodobieństwem około 45% i dopiero ich śmierć przy
braku powstawania nowych terytoriów może doprowadzić do spadku zagęszczenia
populacji. A początkowy spadek tego zagęszczenia wcale nie zmniejsza agresji
samców. Dlatego przy identycznej
liczbie samców zmiany w liczbie skupisk i w wysokości róży idą w odwrotnym
kierunku w zależności od fazy cyklu – wzrastającej lub malejącej. Zmiany zachowań samców możliwe są dopiero przy bardzo niskim ich
zagęszczeniu. Widać to wyraźnie na obu wykresach ryc. 4.4.
Wyjaśnienie cykli
populacyjnych pardwy tak w oparciu o model teoretyczny, jak i dane
empiryczne wydaje się być ważnym krokiem z zrozumieniu natury takich
cykli. Być może podobne zjawiska
występują w populacjach lemingów, norników i innych ssaków, u których znane
są wyraźne cykliczne zmiany wielkości populacji. Może to być jednak bardzo
trudne do ustalenia, bo ssaki są dużo trudniejsze do obserwacji niż ptaki.
Potwierdza to tezy przedstawione w rozdziale 9, że dla zrozumienia procesów
populacyjnych potrzebna jest znajomość indywidualnych różnic, rozmieszczenia
osobników w przestrzeni i ich relacji z sąsiadami. Takie badania znacznie
łatwiej prowadzić na ptakach, niż na ssakach. Być może w przyszłości uda się
także potwierdzić na gryzoniach opisany dla pardwy szkockiej mechanizm
powstawania cykli populacyjnych.
Ryc.
4.4. Zależności między liczbą terytorialnych samców, liczbą grup
krewniaczych, każda składająca się z co najmniej dwóch sąsiadujących
terytoriów (góry wykres), oraz wysokością róży będącą wskaźnikiem agresji
miedzy samcami (dolny wykres) w latach od 1994 do 2002. Wykresy te pokazują
mechanizm opóźnienia w reakcjach między tymi trzema czynnikami. Aby populacja
rosła potrzebne są osobniki spokrewnione w sąsiedztwie i niski poziom
agresji. Taki proces wymaga czasu, tak że populacja osiąga krytyczne
zagęszczenie po kilku latach, a gdy już osiągnie to liczba grup krewniaczych
spada i agresja rośnie, a w konsekwencji spada zagęszczenie. Ale nawet gdy
spadnie do takiego poziomu, który w ubiegłych latach umożliwiał wzrost, to
jednak przy spadku zagęszczenia kilka lat musi minąć zanim odbuduje się
struktura pokrewieństwa i spadnie agresja. Dlatego przy tym samym poziomie
zagęszczenia populacji przestrzenna struktura pokrewieństwa i poziom agresji
różnią się w zależności od tego czy mamy do czynienia ze wzrostem czy
spadkiem tego zagęszczenia. Dalsze objaśnienia w tekście. Rycina wg Piertney et al (2008) zmodyfikowana. Hendry R., Bacon P. J., Moss R., Palmer S. C. F., McGlade J. 1997, A two-dimensional individual-based model
of territorial behaviour: possible population consequences of kinship in red
grouse. Ecological Modelling 105:
23-39. Piertney
S. B., Lambin X., Maccoll
A. D. C., Lock K., Bacon P. J., Dallas J. F., Leckie
F., Mougeot F., Racey
P. A., Redpath S., Moss R. 2008. Temporal changes
in kin structure through a population cycle in a territorial bird, the red
grouse Lagopus lagopus scoticus. Molecular Evolution 17: 2544-2551. Strona 215, wiersz 6 od dołu: tytuł pisma Animal Behaviour daj kursywą Animal Behaviour Strona 218, wiersz 15 od dołu: zamiast i zebra daj
in zebra Strona 219, wiersz 10 do dołu: słowo sociaux daj duzą
literą Sociaux |
|||